






La neurobiología y la patofisiología del insomnio

INTRODUCCIÓN
Esta revisión provee percepciones nuevas del sistema sueño y vigilia y cubre revisiones recientes acerca de la nosología y el diagnóstico del insomnio, los avance en el entendimiento de que el insomnio puede ocurrir como resultado de una hiperexcitación, y cómo el sueño y la excitación están regulados por neurotransmisores clásicos y por neuropéptidos conocidos como orexina A y B (también conocidos como hipocretina 1 y 2).
No hay una definición simple que abarque todo lo que es el sueño. Los hallazgos de esta investigación proveen el entendimiento de que es un estado neurobiológico complejo.
NOSOLOGÍA DEL INSOMNIO
En 2013, cónsono con el incremento en el entendimiento de los desórdenes del sueño y vigilia, la epidemiología, la patofisiología y las investigaciones genéticas que han ocurrido desde la publicación de la 4ta Edición del Manual de Diagnóstico y Estadísticas de Desórdenes Mentales® (DSM-IV), la Asociación Americana de Psiquiatría actualizó la nosología y los criterios para el diagnóstico asociados al insomnio en la 5ta Edición del Manual de Diagnóstico y Estadísticas de los Trastornos Mentales® (DSM-V2). Los términos insomnio primario e insomnio secundario fueron remplazados por el término general trastornos del insomnio. A pesar de que la terminología ha cambiado, se entiende que el insomnio ocurre a menudo junto con un trastorno mental o médico u otro trastornos del sueño1.
La epidemiología y las comorbilidades relacionadas al insomnio
El insomnio es el trastorno del sueño más común. Los estudios han arrojado tasas de prevalencia bastante consecuentes que fluctúan entre un 4% a un 24% en la población general. Se debe observar que las tasas de prevalencia informadas varían según los criterios diagnósticos utilizados, o si se considera el insomnio informado por la propia persona. Además, las investigaciones epidemiológicas muestran una variabilidad significativa en las tasas de prevalencia como resultado de las distintas técnicas de medición y las poblaciones estudiadas16.
Hay un número de factores que se asocian con un aumento en el riesgo del insomnio: Uno es la edad; las personas de mayor edad están más propensas al insomnio. Las mujeres están más propensas a experimentar insomnio que los hombres. Y hay una agregación familiar aparente que se presume que es debido a características genéticas que corren en las familias. Los trastornos de sueño y vigilia, en general, ocurren con mucho más frecuencia en pacientes con trastornos psiquiátricos en comparación con la población general, según se evidencia por una tasa de prevalencia de aproximadamente un 40%. En el STAR* D se encontró que el 84.7% de las personas diagnosticadas con un trastorno de depresión mayor (DDM) no psicótico muestra características clínicas de insomnio (Sunderajan et al., 2010). El insomnio ocurre en aproximadamente un 24% de los pacientes con trastornos de ansiedad y también puede ser comórbido, pero a una incidencia menor, con distimia y abuso de alcohol y drogas8. Para los psiquiatras y otros profesionales de la salud que atienden pacientes con trastornos psiquiátricos, es importante reconocer que el insomnio es común en ellos7.
El insomnio puede afectar muchos aspectos de la vida de una persona. El insomnio puede afectar la concentración y la memoria, aumentar el riesgo de depresión incidental o la recurrencia de un episodio depresivo, disminuir la habilidad del paciente para disfrutar las relaciones de familia y sociales y aumentar el riesgo de caídas y de accidentes de vehículos de motor2.
Las condiciones médicas, como la apnea del sueño, el dolor crónico, la enfermedad pulmonar obstructiva crónica, la hipertensión, la diabetes y la artritis se pueden exacerbar o presentarse junto con el insomnio y, también, el insomnio puede empeorar estas condiciones (p.ej., apneas prolongadas o arritmias electrocardiográficas durante el sueño REM o despertarse confundido con demencia)12. Es imperativo que los médicos que tratan las quejas de sueño y vigilia investíguen la presencia de comorbilidades. Específicamente, es más probable que los trastornos del sueño aparezcan antes de la aparición del DDM. Su presencia y persistencia predicen episodios futuros de DDM y predicen la recurrencia y un resultado peor en pacientes con DDM (i.e., cronicidad y suicidalidad)28. De manera similar, una reducción del sueño a menudo precede el inicio de manía en el desorden bipolar. Hay evidencia de la importancia de reconocer y tratar el insomnio para prevenir o mitigar el DDM y su recurrencia6.
Los pacientes con insomnio a menudo expresan su experiencia con el insomnio como una aceleración del cerebro que no puede parar. Esto puede causar mucha angustia y causar problemas médicos y psiquiátricos. Los médicos deben explicar a los pacientes que el insomnio puede ser un trastorno asociado a una hiperexcitación excesiva en lugar de una disfunción del sistema del sueño. Esta hiperactividad es demostrada por un número de mediciones fisiológicas, como aumento en la frecuencia cardiaca, aumento del ritmo metabólico y aumento de la temperatura corporal y disminución en la habilidad para dormir de día o de noche. La etiología de esta hiperactividad se desconoce; sin embargo, hay la hipótesis de que puede ocurrir relacionado a un cambio en el equilibrio entre las vías que activan el sueño y las vías que activan la vigilia3.
La neurobiología del sueño y la vigilia _ Los circuitos neurales que activan la vigilia surgen de grupos de células en el tallo cerebral, el hipotálamo y prosencéfalo que activan la corteza cerebral (Fig. 1). Históricamente, las vías neurales que emergen del tallo cerebral se conocen como el sistema activador reticular ascendente, pero en los pasados 25 años, los investigadores han establecido que esta vía está compuesta de varias proyecciones específicas que liberan los neurotransmisores acetilcolina y monoaminas, como la norepinefrina, la dopamina, la serotonina y la histamina19. Estos grupos de células incluyen neuronas histaminérgicas en el núcleo tuberomamilar (NTM), las neuronas serotoninérgicas del núcleo del rafe y las neuronas noradrenérgicas del locus coeruleus (LC). Cada uno de estos grupos parece influenciar distintos aspectos de la excitación, y actúan de forma colectiva para producir una excitación total en condiciones adecuadas. La importancia clínica de estas vías queda demostrada por la sedación mostrada por los antagonistas de las mono aminas y los anticolinérgicos (como el haloperidol, la difenhidramina y la mirtazapina)20.
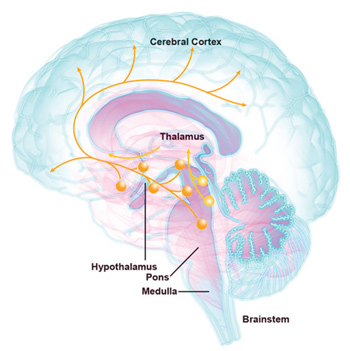
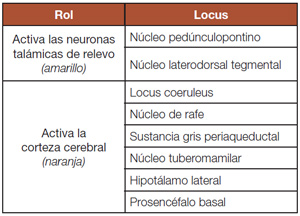 Las neuronas orexinérgicas son otro elemento clave del sistema activador de la vigilia20. La orexina A y la orexina B (también conocidas como hipocretina 1 y 2) son neurotransmisores peptídicos que se enlazan a los receptores OX1 y OX2, donde tienen efectos excitatorios.
Las neuronas orexinérgicas son otro elemento clave del sistema activador de la vigilia20. La orexina A y la orexina B (también conocidas como hipocretina 1 y 2) son neurotransmisores peptídicos que se enlazan a los receptores OX1 y OX2, donde tienen efectos excitatorios.
A diferencia de otros neurotransmisores activadores de la vigilia que son producidos por una variedad de núcleos, las orexinas son producidas solamente por un pequeño grupo de neuronas en el hipotálamo lateral. Similar a los otros sistemas activadores de la vigilia, las orexinas se liberan durante la vigilia, y estas excitan una amplia variedad de regiones cerebrales, con efectos especialmente potentes en las otras regiones cerebrales activadoras de la vigilia. Una inyección única de orexina-A en el sistema nervioso central de una rata puede activar la vigilia por varias horas. La importancia del sistema es bastante evidente en la narcolepsia, en la que la pérdida crónica y selectiva de neuronas productoras de orexina induce una condición clínica caracterizada por transiciones descontroladas y súbitas hacia el sueño5.
Las orexinas también activan otros aspectos de la excitación. Las orexinas aumentan la actividad en las regiones del cerebro que regulan la motivación y la recompensa, y los antagonistas de orexina pueden aliviar la urgencia de drogas en animales experimentales. Las orexinas también aumentan la frecuencia cardiaca, la presión arterial y la actividad locomotora. La integración de estos aspectos variados de la excitación por las orexinas pueden ayudar a asegurar que una persona se mantenga alerta, activa y motivada a las horas correctas del día20. Durante el sueño no REM y REM, las neuronas productoras de orexina están inhibidas por neuronas que liberan ácido γ-aminobutírico (GABA) y galanina. El núcleo preóptico ventrolateral (POVL) es el mejor estudiado de estos sistemas GABAérgicos, y una lesión en este núcleo induce cantidades sustancialmente reducidas de sueño REM y no REM. (Fig. 2)19.
Por lo tanto, contrario a la redundancia en la señalización de la vigilia en el cerebro, se han identificado pocas poblaciones neurales activadoras del sueño.
El sueño REM se caracteriza por movimientos oculares rápidos, atonía muscular y producción de sueños. Durante el sueño REM, las neuronas monoaminérgicas se hiperpolarizan, pero una acumulación de neuronas colinérgicas en la protuberancia (núcleo tegmental laterodorsal y pedunculopontino) se despolarizan. Estas y otras neuronas pontinas juegan roles esenciales en la producción de la activación cortical y la atonía muscular típicas del sueño REM. Estas vías activadoras del sueño REM están inhibidas fuertemente por los neurotransmisores mono amina, y en la práctica clínica, el sueño REM a menudo se suprime mediante fármacos que aumentan la serotonina o la norepinefrina, como los inhibidores selectivos de la recaptación de serotonina y los inhibidores duales de la recaptación de serotonina y norepinefrina14.
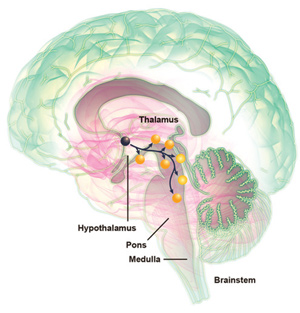
 Las orexinas también regulan el sueño REM. Durante la vigilia, la orexina activa las neuronas mono aminérgicas y otras regiones del tallo cerebral que suprimen el sueño REM. Sin embargo, con la pérdida crónica de neuronas orexina en la narcolepsia, el sueño REM puede ocurrir en cualquier momento del día, y los elementos del sueño REM se mezclan con el estado de vigilia. Por ejemplo, pueden ocurrir sueños REM al irse a dormir o al despertar, y esto se puede mezclar con la vigilia, causando alucinaciones hipnagógicas. Aún más, puede ocurrir parálisis del sueño REM al despertar, o aún en medio de la vigilia, y manifestarse como parálisis del sueño o cataplexia, respectivamente5.
Las orexinas también regulan el sueño REM. Durante la vigilia, la orexina activa las neuronas mono aminérgicas y otras regiones del tallo cerebral que suprimen el sueño REM. Sin embargo, con la pérdida crónica de neuronas orexina en la narcolepsia, el sueño REM puede ocurrir en cualquier momento del día, y los elementos del sueño REM se mezclan con el estado de vigilia. Por ejemplo, pueden ocurrir sueños REM al irse a dormir o al despertar, y esto se puede mezclar con la vigilia, causando alucinaciones hipnagógicas. Aún más, puede ocurrir parálisis del sueño REM al despertar, o aún en medio de la vigilia, y manifestarse como parálisis del sueño o cataplexia, respectivamente5.
La regulación de las transiciones entre la vigilia y el sueño _ La inhibición mutua entre los sistemas activadores del sueño y los sistemas activadores de la vigilia, ayudan a producir una vigilia total y un sueño total. El GABA del núcleo POVL inhibe los sistemas activadores de la vigilia, y las mono aminas y la acetilcolina inhiben el núcleo POVL. Por lo tanto, durante la vigilia, los neurotransmisores inhiben el núcleo POVL, eliminando cualquier interferencia del POVL en este estado, y permite que los sistemas estén totalmente activos y alertas. Lo opuesto ocurre durante el sueño, cuando las neuronas POVL se liberan desde cualquier inhibición de los sistemas activadores de la excitación.
Algunos investigadores se refieren a esto como un interruptor neural tipo flip-flop porque, al igual que un circuito eléctrico del mismo nombre, la inhibición mutua de los sistemas activadores del sueño y la vigilia asegura la excitación total o el sueño total, con poco tiempo entre los estados intermedios20. Una desventaja de un interruptor flip-flop es que puede resultar en transiciones indeseadas con poco aviso. Sería ventajoso desde el punto de vista biológico, tener un mecanismo que estabilice el interruptor, y asegurar que ocurra un sueño adecuado. Los neuropéptidos orexinérgicos, descubiertos en 1998, parecen tener una función importante en esta estabilización. (Fig. 3)18.

¿Cuál es la función de las vías orexinérgicas?
Este cambio entre la vigilia y el sueño es estabilizado por los neuropéptidos orexinérgicos. Principalmente, esto ocurre mediante los efectos excitatorios directos de las orexinas en otros grupos celulares activadores de la vigilia27.
Aún más, las neuronas orexinérgicas son autoexcitatorias; la exocitosis de las orexinas induce una liberación local de glutamato y como consecuencia aumenta la actividad de las neuronas orexinérgicas. Los investigadores entienden que esta asa de retroalimentación positiva puede llevar a una actividad sostenida de las neuronas orexinérgicas la cual luego provoca una actividad sostenida en los otros sistemas de excitación para producir periodos prolongados de vigilia. Por el contrario, la pérdida de las neuronas orexina dificulta a los pacientes o animales con narcolepsia a mantener periodos extendidos de vigilia11.
Diferencias en la distribución y los efectos del GABA y la orexina. _ Es importante diferenciar la distribución y los efectos del GABA y las orexinas en el cerebro. GABA es el neurotransmisor inhibitorio principal del sistema nervioso central y los receptores de GABA se encuentran en todo el cerebro24 mientras que la distribución de los receptores de orexina es más limitada. Hay áreas del cerebro que no tienen inervación orexinérgica, como el cerebelo, donde no hay sinapsis ni receptores orexinérgicos. Los efectos principales del sistema de orexinas son ayudar a sostener periodos prolongados de vigilia, activar la vigilia y suprimir el sueño REM. Los otros efectos de la orexina son menores y solo tienen efectos mínimos en el tono autonómico y en la respiración20.
La hiperexcitación como el aspecto principal del insomnio
Evidencia fisiológica _ Las investigaciones han demostrado que el insomnio está fuertemente relacionado a la hiperactividad en los sistemas neurales de la excitación. La hiper-excitación entre los sujetos con insomnio se evidencia por la elevación de la temperatura corporal, la frecuencia cardiaca, los niveles de catecolaminas, la respuesta al estrés y otros cambios fisiológicos en comparación con sujetos control sanos. Estas elevaciones en las mediciones fisiológicas sustentan la conceptualización del insomnio como un trastorno de hiperexcitación4. Esta hiperexcitación no está limitada al sueño y está presente durante el día también. El nivel de excitación diurno puede medirse mediante la Prueba de Latencia Múltiple del Sueño (PLMS) [Multiple Sleep Latency Test (MSLT)], en la cual a los sujetos se les da cinco oportunidades de tomar una siesta durante el día. Las latencias de sueño cortas de esta prueba indican somnolencia diurna, mientras que latencias más largas indican niveles más altos de excitación. Entre los pacientes con insomnio, los más alerta (es decir, con la latencia del sueño más larga), tienen el peor sueño durante la noche. Aquellos con insomnio que están muy alertas (es decir, tienen latencias de sueño de 15-18 minutos) solo duermen alrededor de 6 horas en la noche. Los resultados de este estudio indican que las latencias del sueño prolongadas en la MSLT se correlacionan negativamente con el tiempo del sueño en pacientes con insomnio. Esto es lo opuesto de lo observado en voluntarios sanos normales, en que mientras menos se duerme o mientras más severo el disturbio del sueño, mayor el grado de somnolencia diurna subjetiva. Por tanto, menos sueño está relacionado a una disminución en la alerta en la población general, pero una reducción del sueño en el insomnio está relacionado con un aumento en la alerta17.
Los niveles de norepinefrina son más altos en algunos pacientes con insomnio. Los pacientes con insomnio con latencias del sueño más prolongadas en la MSLT tienen niveles de norepinefrina más altos que los pacientes con latencias del sueño más cortas en la MSLT. Aún más, los niveles de norepi-nefrina se encontraron significativamente elevados en pacientes con insomnio respecto de los sujetos control. Por lo tanto, hay sincronía entre niveles elevados de norepinefrina, aumento en la alerta diurna y disminución total del tiempo del sueño25.
Las personas con insomnio tienen niveles elevados de cortisol respecto de los controles, lo cual es más notable alrededor del inicio del sueño, contribuyendo a evidenciar que el insomnio es un disturbio de hiperexcitación. De forma interesante, los niveles de cortisol en sujetos con y sin trastornos del sueño se diferencian principalmente antes del inicio del sueño, momento en el cual pensar acerca de irse a la cama puede actuar con un factor estresante y aumentar los niveles de cortisol para la persona con insomnio26.
En un estudio aparte de pacientes con insomnio, los investigadores monitorizaron la frecuencia cardiaca al colocar las manos de los sujetos en un cubo de agua fría (un estresante) cuando estaban despiertos, y la frecuencia cardiaca aumentó en los que tenían insomnio en comparación con los sujetos control23. Todos estos hallazgos son cónsonos con el insomnio como un trastorno de hiperexcitación, contrario a uno de disfunción del sistema del sueño.
Las imágenes como evidencia _ También, se ha documentado la evidencia de hiper-excitación en pacientes con insomnio en los estudios de imágenes. Por lo general, el insomnio está relacionado a un aumento en el metabolismo cerebral. Al comparar los datos de sujetos con y sin insomnio, las tomografías de emisión de positrones (PET) demostraron un aumento en el metabolismo de la glucosa cerebral en pacientes con insomnio durante el sueño y en vigilia, una disminución menor en el metabolismo en la transición de vigilia a sueño en las regiones activadoras de la vigilia, y una reducción del metabolismo en la corteza prefrontal en vigilia. Los centros de excitación, incluidos el mesencéfalo y el hipotálamo, muestran menor reducción en la actividad durante el sueño en pacientes con insomnio. Es extremadamente importante entender que estos pacientes con insomnio estaban durmiendo cuando se tomaron las imágenes, lo que muestra que las personas con insomnio tienen un aumento en el metabolismo cerebral durante el sueño no REM. Esta actividad puede observarse con un aumento total del metabolismo cerebral a lo largo de las 24 horas del día15. Este aumento en el metabolismo cerebral provee más evidencia de una hiperexcitación en pacientes con insomnio.
CONCLUSIONES
El insomnio puede causar problemas médicos y psiquiátricos significativos. Las investigaciones clínicas proveen evidencia de que el insomnio es un trastorno de hiperexcitación evidenciado mediante aumentos en la frecuencia cardiaca, el índice metabólico, una actividad rápida del EEG, los niveles de catecolaminas y aumento en la reactividad al estrés. La evidencia actual sugiere que el insomnio puede ser un desorden de excitación excesiva en lugar de un desorden del sistema del sueño. Las investigaciones básicas han demostrado que el POVL usa GABA para inhibir los sistemas de excitación, y los sistemas de excitación usan mono aminas y acetilcolina para inhibir el POVL. La inervación orexinérgica, contrario a GABA, no ocurre en todas las áreas del cerebro. Aun dentro de las áreas que las neuronas orexinérgicas sí inervan, la orexina es más selectiva hacia grupos celulares específicos. Los neuropéptidos orexinérgicos excitan las otras regiones cerebrales activadoras de la vigilia para sostener la vigilia y la vigilancia. Aunque queda mucho por aprender acerca de la neurobiología subyacente del insomnio clínico, es posible que la hiperexcitación y el mal dormir sean debido a una hiperactividad de los sistemas activadores de la vigilia.
Conflictos de interés Luis Mejía pertenece al Departamento Médico de Merck, Puerto Rico.
Referencias -# American Psychiatric Association. (2000). Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition, Text Revision®, (Copyright ©2000). Washington, DC: American Psychiatric Association. -# American Psychiatric Association. (2013). Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition®, (Copyright ©2013). Arlington, VA: American Psychiatric Publishing. -# Alexandre, Andermann and Scammell. Current Opinion in Neurobiology (2013), 23:752– 759. -# Bonnet, M.H., Arand, D.L., Hyperarousal and insomnia: state of science. Sleep Medicine Reviews (2010). 14, 9–15. -# Burgess, C. R., & Scammell, T. E. (2012). Narcolepsy: neural mechanisms of sleepiness and cataplexy. J Neurosci, 32(36), 12305-12311. -# Chellappa, S., & Araújo, J. (2007). Sleep disorders and suicidal ideation in patients with depressive disorders. Psychiatry Res, 153, 131-136. -# Drake, C., Richardson, G., Roehrs, T., Scofield, H., & Roth, T. (2004). Vulnerability to stress-related sleep disturbance and hyperarousal. Sleep, 27(2), 2 8 5 -291. -# Ford, D. E., & Kamerow, D. B. (1989). Epidemiologic study of sleep disturbances and psychiatric disorders. An opportunity for prevention? JAMA, 262(11), 1479-1484. -# Franzen, P. L., & Buysse, D. J. (2008). Sleep disturbances and depression: risk relationships for subsequent depression and therapeutic implications. Dialogues Clin Neurosci, 10(4), 473-481. -# Jeffrey M. Ellenbogen & Edward F. Pace-Schott. Drug-induced sleep: theoretical and practical considerations. Eur J Physo (2012) 463:177–186. -# Kumar, S, Sagili, H. Journal of clinical and diagnostic research: JCDR Etiopathogenesis and neurobiology of narcolepsy: a review. Volume: 8 Issue: 2 Pages: 190-5 (2014). -# Kraus Stefanie, Rabin Laura. Sleep America: Managing the crisis of adult chronic insomnia and associated conditions. Journal of Affective Disorders 138 (2012) 192–212. -# Morin M Charles, Benca Ruth. Chronic insomnia. Lancet (2012); 379: 1129–41. -# Nishino, S., & Mignot, E. (1997). Pharmacological aspects of human and canine narcolepsy. Prog Neurobiol, 52(1), 27-78. -# Nofzinger, E. A., Buysse, D. J., Germain, A., Price, J. C., Miewald, J. M., & Kupfer, D. J. (2004). Functional neuroimaging evidence for hyperarousal in insomnia. Am J Psychiatry, 161(11), 2126-2128. -# Ohayon, M. M., & Roth, T. (2001). What are the contributing factors for insomnia in the general population? J Psychosom Res, 51(6), 745-755. -# Roehrs, T. A., Randall, S., Harris, E., Maan, R., & Roth, T. (2011). MSLT in primary insomnia: stability and relation to nocturnal sleep. Sleep, 34(12), 1647-1652. -# Sakurai T. Current Opinion in Neurobiolg., Vol 23 (5), (2013), 76–766. -# Saper, C. B., Scammell, T. E., & Lu, J. (2005). Hypothalamic regulation of sleep and circadian rhythms. Nature, 437(7063), 1257-1263. -# Scammell, T. E., & Winrow, C. J. (2011). Orexin receptors: pharmacology and therapeutic opportunities. Annu Rev Pharmacol Toxicol, 51, 243-266. -# Sunderajan, P et al. (2010). Insomnia in patients with depression: a STAR*D report. CNS Spectr, 15(6), 394-404. -# Steiner, Michel A; Winrow, Christopher J. Frontiers in neuroscience Volume: 8 Pages: 158 (2014). -# Stepanski, E., Glinn, M., Zorick, F., Roehrs, T., & Roth, T. (1994). Heart rate changes in chronic insomnia. Stress Med, 10(4), 261-266. -# Uusi-Oukari M, Korpi ER Regulation of GABA(A) receptor subunit expression by pharmacological agents. Pharmacol Rev (2010) 62:97–135. -# Vgontzas, A. N., Bixler, E. O., Lin, H. M., Prolo, P., Mastorakos, G., Vela-Bueno, A.,Chrousos, G. P. (2001). Chronic insomnia is associated with nyctohemeral activation of the hypothalamic-pituitary-adrenal axis: clinical implications. J Clin Endocrinol Metab, 86(8), 3787-3794. -# Vgontzas, A. N., Tsigos, C., Bixler, E. O., Stratakis, C. A., Zachman, K., Kales, A., Chrousos, G. P. (1998). Chronic insomnia and activity of the stress system: a preliminary study. J Psychosom Res, 45(1), 21-31. -# Winrow CJ, Gotter AL, Cox CD, Doran SM, Tannenbaum PL, Breslin MJ, Garson SL, Fox SV, Harrell CM, Stevens J, Reiss DR, Cui D, Coleman PJ, Renger JJ (2011) Promotion of sleep by suvorexant-a novel dual orexin receptor antagonist. J Neurogenet 25:52–61. -# Wojnar, M.,et al. (2009). Sleep problems and suicidality in the National Comorbidity Survey Replication. J Psychiatr Res, 43(5).
Agradecimiento especial: Al Dr. Christopher J. Winrow, Director, Descubrimiento Molecular, Merck; al Dr. Felipe Arbelaez, Director Médico de Merck, PR por las mejoras a la revisión.






